内容
P2X2は、細胞外のATPによって活性化される陽イオンチャネルである。P2X2は、2回膜貫通型のサブユニット3個が会合して形成されるが、その分子内に、典型的な膜電位センサーと考えられる部位は存在しない。我々は、これまでの研究で、P2X2が膜電位センサー部位を有しないにも関わらず、ATP投与後の定常状態で膜電位依存性のゲーティングを示すことを明らかにした。
近年、Eric Goueuxの研究グループによりゼブラフィッシュのP2X4のATP 非結合時(閉状態)およびATP結合時(開状態)の結晶構造が解かれ、ATP結合に伴う構造変化が示された。しかし、結晶構造には、膜電位やその変化は反映されていないので、膜電位依存的にどのような構造変化が起こるかについては、未だ情報は無い。
本研究では、膜電位とATPに依存するラットP2X2の構造の変化を、「点変異により導入したCys残基のCys修飾剤による修飾の速度の、状態に依存する違いとして捉える」ことを目的とした。そこで、ATP結合部位とチャネルポアを連結するリンカー部位に位置するAsp315とIle67に点変異によりCys残基を導入したコンストラクト (D315C & I67C) を作成しツメガエル卵母細胞に発現させた。そして、二本刺し膜電位固定法により、Cys残基に結合するCd2+ の投与によるチャネル電流の変化を記録し、その速度を、異なる状態間で比較解析した。
ATP存在下において、過分極状態と脱分極状態での速度を比較したところ、過分極状態の方が、優位に修飾が速かった(図1)。また、修飾速度は、ATP濃度の上昇につれて加速した(図1)。次に、ATP非存在下と存在下での修飾速度を比較したところ、ATP非存在下では非常に遅く、過分極状態でも、脱分極状態でも、ほとんど修飾が起らなかった。これらの結果から、リンカー部位において、膜電位とATP結合の有無に依存して構造の変化が起きることが明らかになった。
さらに、膜電位依存性のゲーティングを示さないチャネルポアの変異体Thr339Ser (T339S) を対象に、同様な実験を行ったところ、Cys変異体 (D315C&I67C&T339S) のCd2+ による修飾速度は、過分極と脱分極で差を示さなかった(図2)。この結果は、捉えた膜電位に依存する構造変化が、膜電位依存的なポアのゲーティングに伴うものであることを示している。
本研究によって、膜電位センサーを有しないP2X2のATP結合部位とチャネルポアをつなぐリンカー領域において、ATP結合に依存して、また膜電位に依存して、構造変化が起こることが、点変異により導入したCys残基のCd2+ による修飾速度の違いとして捉えられた。この結果から、P2X2が、ATPと膜電位の両方を複合的に感知して構造変化を起こすチャネルであることが示された(図3)。
図1
Image may be NSFW.Clik here to view.
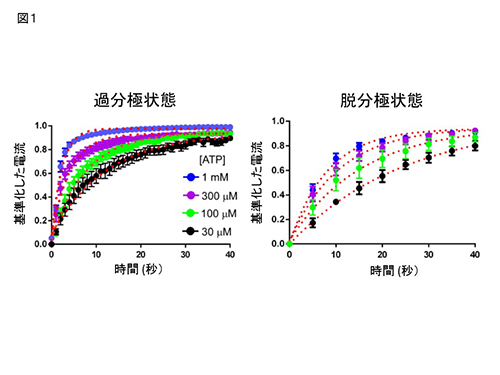 Cys変異体 (D315C & I67C) の、Cd2+ 投与による電流修飾速度は、膜電位およびATP濃度に依存して変化する。
Cys変異体 (D315C & I67C) の、Cd2+ 投与による電流修飾速度は、膜電位およびATP濃度に依存して変化する。図2
Image may be NSFW.
Clik here to view.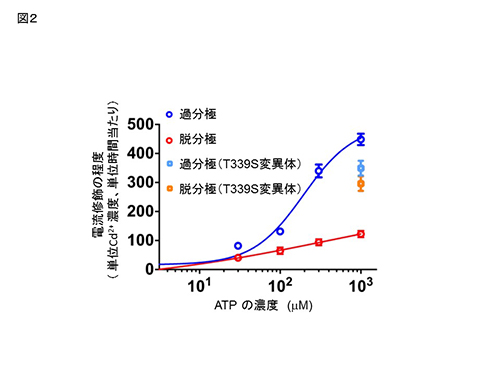 膜電位依存的ゲーティングを失ったT339S変異体 (D315C&I67C&T339S) の、Cd2+ による修飾の速度は、膜電位に依存して変化しない。青、赤は、図1で示したCys変異体 (D315C & I67C)のデータ。
膜電位依存的ゲーティングを失ったT339S変異体 (D315C&I67C&T339S) の、Cd2+ による修飾の速度は、膜電位に依存して変化しない。青、赤は、図1で示したCys変異体 (D315C & I67C)のデータ。
図3
Image may be NSFW.
Clik here to view.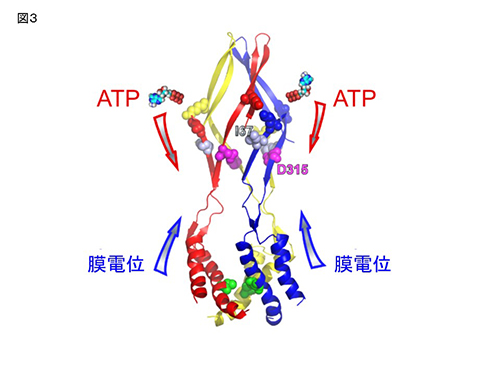 P2X2のチャネルポアとATP結合部位をつなぐリンカー領域において、ATP結合と膜電位の両方に依存して構造変化が起こる。
P2X2のチャネルポアとATP結合部位をつなぐリンカー領域において、ATP結合と膜電位の両方に依存して構造変化が起こる。
論文情報
Batu Keceli and Yoshihiro Kubo
J Physiol September 2014; doi:10.1113/jphysiol.2014.278507
